| TOP>梈崌尋媶僐乕僗 暯惉17擭搙丂尋媶曬崘 | |
帀朾巁庴梕懱俧俹俼侾俀侽偺僎僲儉栻棟妛揑夝愅僎僲儉憂栻壢妛暘栰丂栴栰丂晲柧 尋 媶 巜 摫 庡 擟 丗 僎僲儉憂栻壢妛暘栰丂嫵庼丂捯杮丂崑嶰 尋媶巜摫嫤椡幰丗惗懱忣曬惂屼妛暘栰丂嫵庼丂拞嶳丂榓媣 尋媶撪梕梫巪丂俧俹俼120偺儕僈儞僪偱偁傞帀朾巁偺僇儖儃僉僔儖婎偑庴梕懱偺嵶朎奜儖乕僾偵懚嵼偡傞墫婎惈傾儈僲巁巆婎偲憡屳嶌梡偡傞偲壖掕偟丄偙傟傜係儢強偺傾儈僲巁巆婎傪揰曄堎摫擖偵傛傝傾儔僯儞偵抲姺偟偨丅嶌惉偟偨曄堎懱傪偦傟偧傟俫俤俲293嵶朎偵堦夁惈偵敪尰偝偣偨屻丄儕僈儞僪偱偁傞α-儕僲儗儞巁巋寖偵敽偆俤俼俲偺妶惈壔傪應掕偟偨丅偦偺寢壥丄戞堦嵶朎奜儖乕僾偵懚嵼偡傞俼99俙曄堎懱偺傒偵偍偄偰尠挊側俤俼俲偺儕儞巁壔梷惂偑妋擣偝傟偨丅偦偺屻丄Tet-On 僔僗僥儉偵傛傝僪僉僔僒僀僋儕儞偱敪尰偑桿摫偝傟傞埨掕敪尰嵶朎姅傪嶌惉偟丄帀朾巁偍傛傃揤慠暔傛傝拪弌偝傟偨儕僈儞僪偵懳偟偰俤俼俲偺儕儞巁壔偲嵶朎撪俠倎2+偺曄摦傪應掕偟偨寢壥丄帀朾巁偺儕僈儞僪偼俼99俙曄堎懱偵偍偄偰尠挊側僔僌僫儖偺梷惂偑擣傔傜傟偨丅堦曽丄帀朾巁偲偼峔憿偺堎側傞揤慠暔傛傝拪弌偝傟偨儕僈儞僪偵偍偄偰偼丄俼99俙偵偍偗傞僔僌僫儖偺梷惂偼擣傔傜傟側偐偭偨丅埲忋偺寢壥偐傜丄帀朾巁偺儕僈儞僪偼俼99俙傪夘偟偰俧俹俼120偲憡屳嶌梡偟偰偄傞帠傪柧傜偐偵偟偨丅 攚宨丂俧抈敀幙嫟栶宆庴梕懱乮俧-protein-coupled receptor丟俧俹俠俼乯偼嵶朎忣曬揱払宯偵偍偄偰庡梫側拠夘場巕偱偁傝丄幘昦偲偺娭學傕怺偔丄憂栻尋媶忋廳梫側昗揑暘巕偱偁傞丅俧俹俠俼拞偵偼儕僈儞僪偑枹抦偺丄偄傢備傞僆乕僼傽儞庴梕懱傕懡悢娷傑傟丄僆乕僼傽儞庴梕懱偺儕僈儞僪扵嶕偍傛傃婡擻夝愅偼嬌傔偰廳梫偱偁傞丅摉尋媶幒偱偼丄偙偺僆乕僼傽儞庴梕懱偺堦偮偱偁偭偨俧俹俼120偺儕僈儞僪偑晄朞榓挿嵔帀朾巁偱偁傞偙偲傪柧傜偐偵偟偨丅俧俹俼120偼挵娗摿堎揑偵敪尰偟偰偍傝丄儅僂僗桼棃偺挵撪暘斿嵶朎俽俿俠-1偵傕敪尰偟偰偄傞丅俽俿俠-1嵶朎偼梀棧帀朾巁偵斀墳偟儁僾僠僪儂儖儌儞傪梀棧偡傞偙偲偑曬崘偝傟偰偍傝丄梀棧帀朾巁巋寖偵傛傞 glucagons like peptide-1乮俧俴俹-1乯偺暘斿偑丄俧俹俼120傪夘偟偰偄傞偙偲偑柧傜偐偲側偭偨丅怽惪幰傜偵傛傞愭偺曬崘偵偍偄偰俽俿俠-1偱偺梀棧帀朾巁巋寖偵傛傞傾億僩乕僔僗梷惂偑 栚揑丂帀朾巁庴梕懱俧俹俼120巋寖偵傛傞俧俴俹-1暘斿婡峔偺慡梕傪夝柧偡傞偨傔丄儕僈儞僪偲庴梕懱偺捈愙揑側憡屳嶌梡傪徹柧偡傞偲摨帪偵丄庴梕懱妶惈壔偐傜俧俴俹-1偺暘斿傑偱偺宱楬傪柧傜偐偵偡傞丅偙傟偵傛傝丄憂栻昗揑暘巕偲偟偰偺俧俹俼120偺惗棟妛揑側婡擻傪柧妋側傕偺偲偡傞偲偲傕偵丄憂栻偵寢傃偮偔曪妵揑側忣曬傪摼傞偙偲傪栚巜偡丅 曽朄丂儕僈儞僪偲庴梕懱偺捈愙揑側憡屳嶌梡傪徹柧偡傞偨傔丄儕僈儞僪偲庴梕懱偺寢崌條幃偺夝愅傪峴偆丅帀朾巁庴梕懱孮偺傾儈僲巁攝楍傾儔僀儞儊儞僩偍傛傃棫懱峔憿偺儌僨儖偐傜儕僈儞僪偲憡屳嶌梡偡傞壜擻惈偺崅偄傾儈僲巁傪慖戰偟丄曄堎摫擖傪峴偆偙偲偱儕僈儞僪寢崌偵昁恵偺傾儈僲巁傪摨掕偡傞丅師偵丄俧俴俹-1偺暘斿婡峔傪夝柧偡傞偨傔丄俧俹俼120偺妶惈壔偵傛傝俧俴俹-1偺暘斿傪婲偙偡俽俿俠-1嵶朎傪儌僨儖偲偟丄僔僌僫儖揱払宯偵懳偡傞栻棟妛揑僣乕儖傪梡偄偨夝愅偍傛傃丄惗壔妛揑丄暘巕惗暔妛揑側夝愅曽朄傪梡偄偰俧俴俹-1暘斿偵娭梌偡傞暘巕偺摨掕傪峴偄丄忣曬揱払宱楬傪柧傜偐偵偡傞丅偝傜偵儕僈儞僪寢崌偺掅壓偟偨揰曄摦摫擖宆偲栰惗宆俧俹俼120庴梕懱偲偺僔僌僫儖揱払宯偺斾妑傪丄摨掕偟偨暘巕庬偵偮偄偰徻嵶偵専摙偡傞偙偲偱丄俧俹俼120偺妶惈壔偐傜俧俴俹-1偺暘斿傑偱偺婡峔傪柧妋偵偡傞丅 寢壥
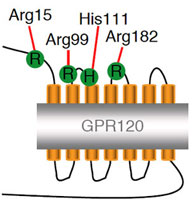 丂 丂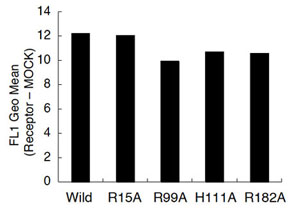 恾侾.墫婎惈傾儈僲巁巆婎偺揰曄堎摫擖偍傛傃敪尰妋擣 帀朾巁偲偺憡屳娭學偵廳梫偱偁傞偲梊應偟偨丄嵶朎奜儖乕僾偵懚嵼偡傞墫婎惈傾儈僲巁巆婎傪傾儔僯儞偵抲姺偟偨乮嵍恾乯丅俥俴俙俧梈崌俧俹俼120傪嶌惢偟偨丅偦傟偧傟俫俤俲293嵶朎偵堦夁惈偵敪尰偝偣丄僼儘乕僒僀僩儊僩儕乕乮俥俙俠俽 俠倎倢倝倐倳倰丗俛俢幮乯偵偰俥俴俙俧峈懱偍傛傃寀岝昗幆僾儘乕僽傪梡偄偰嵶朎枌昞柺偵敪尰偡傞庴梕懱傪應掕偟偨丅廲幉偼奺庴梕懱偲俵俷俠俲偵偍偗傞俧倕倧 俵倕倎値偺嵎暘傪帵偟偨丅
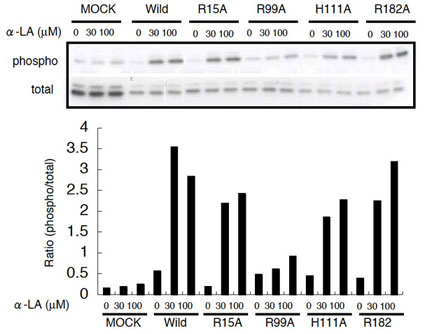
恾俀.俫俤俲293嵶朎偵敪尰偝偣偨俧俹俼120偍傛傃曄堎懱偺俤俼俲妶惈 栰惗宆偍傛傃丄俼15俙丆俫111俙丆俼182俙偺奺曄堎懱傪俫俤俲293嵶朎偵堦夁惈偵敪尰偝偣偨丅偦傟偧傟丄α-俴俙 傪 0丆30丆100 µM 偵偰 5 暘娫巋寖偟偨偺偪丄倫俤俼俲偍傛傃俤俼俲峈懱偵偰専弌偟偨丅壓抜偺朹僌儔僼偼忋抜偺専弌偝傟偨僶儞僪傪悢抣壔偟偨傕偺傪帵偟偨丅
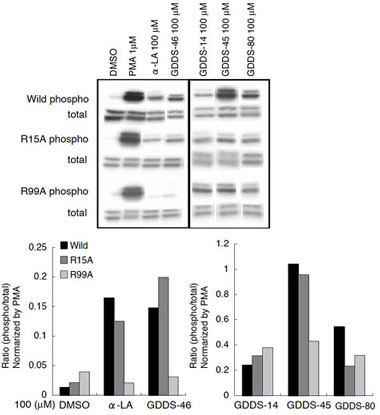
恾俁.埨掕敪尰嵶朎姅偵偍偗傞俧俹俼120偍傛傃曄堎懱偺俤俼俲妶惈 栰惗宆偍傛傃丄俼15俙丆俼99俙偺奺曄堎懱偺僪僉僔僒僀僋儕儞偵偰敪尰傪桿摫偱偒傞埨掕敪尰嵶朎姅乮俫俤俲乯傪嶌惢偟偨丅偦傟偧傟丄僪僉僔僒僀僋儕儞偵偰敪尰傪桿摫偝偣偨屻丄帀朾巁儕僈儞僪偱偁傞 α-俴俙 偍傛傃俧俢俢俽-14丄俧俢俢俽-45丄俧俢俢俽-80 傪 100 µM 偵偰 5 暘娫巋寖偟偨丅偦偺屻倫俤俼俲偍傛傃俤俼俲峈懱偵偰専弌偟偨丅 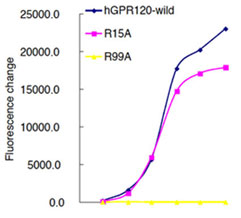 丂 丂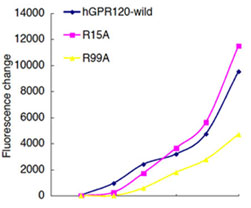 恾係.埨掕敪尰嵶朎姅偵偍偗傞俧俹俼120偍傛傃曄堎懱偺嵶朎撪俠倎2+偺曄摦 栰惗宆偍傛傃丆俼15俙丆俼99俙 偺奺曄堎懱偺僪僉僔僒僀僋儕儞偵偰敪尰傪桿摫偱偒傞埨掕敪尰嵶朎姅乮俫俤俲乯傪嶌惢偟偨丅偦傟偧傟丄僪僉僔僒僀僋儕儞偵偰敪尰傪桿摫偝偣偨偺偪丄奺擹搙偵偍偗傞 α-俴俙 乮嵍恾乯偍傛傃俧俢俢俽-45乮塃恾乯偵偰巋寖偟偨丅 埲忋偺寢壥偐傜丄帀朾巁儕僈儞僪偲俧俹俼120偺憡屳嶌梡偵偼俙倰倗99 偑廳梫偱偁傞偙偲傪帵嵈偟偨丅堦曽丄帀朾巁偲峔憿偺堎側傞儕僈儞僪偵娭偟偰偼丄俙倰倗99 傪夘偝偢憡屳嶌梡偡傞偲峫偊傜傟傞丅 婜懸偝傟傞惉壥丂曄埵摫擖俧俹俼120偺夝愅偵傛傝丄庴梕懱-儕僈儞僪娫偺捈愙揑側憡屳嶌梡傪専徹弌棃傞偼偢偱偁傞丅偝傜偵偙偺寢壥偐傜丄俧俹俼120偵懳偡傞儕僈儞僪寢崌偺棫懱峔憿儌僨儖偺嶌惉偑壜擻偲側傝丄怴婯儕僈儞僪偺扵嶕傊偲栶棫偮偙偲偑婜懸偝傟傞丅偝傜偵丄俧俴俹-1暘斿丄惂屼婡峔偺夝愅偵傛傝丄尞偲側傞忣曬揱払宯偲娭楢偡傞暘巕庬偑柧傜偐偵側傞偱偁傠偆丅偙傟偵傛傝丄俧俹俼120偺挵娗偵偍偗傞惗棟揑婡擻偑偝傜偵柧妋偲側傝丄栻暔奐敪忋丄廳梫側忣曬偑摼傜傟傞偙偲偑婜懸偝傟傞丅挵娗偐傜暘斿偝傟傞俧俴俹-1偼丄僀儞僗儕儞暘斿偺桿摫媦傃僌儖僇僑儞暘斿傪梷惂偟偡傞偙偲偐傜丄俧俴俹-1暘斿丄惂屼婡峔偺暘巕儗儀儖偱偺棟夝偐傜丄摐擜昦丄旍枮丄愛怘挷愡偵懳偡傞怴偨側帯椕昗揑暘巕敪尒偺壜擻惈傕偁傞丅埲忋丄杮尋媶僾儘僕僃僋僩偱偼丄峀偄尋媶暘栰偺楢実偐傜丄帀朾巁庴梕懱俧俹俼120偺儕僈儞僪寢崌夝愅偲丄俧俴俹-1暘斿偵偍偗傞婡擻夝愅偲傪曪妵揑偵峴偄丄僎僲儉憂栻偵傛傞怴婯偺憂栻昗揑暘巕偺摨掕偐傜丄憂栻墳梡傊偲敪揥偡傞尋媶偺払惉偑婜懸偝傟傞丅 |
|